Il daltonismo consiste nell'incapacità di percepire in modo corretto alcuni colori, come il rosso dal verde; essendo i geni che servono per codificare questi colori sullo stesso cromosoma, se uno non funzione, vedo solamente l'altro colore, e viceversa.

L'emofilia
consiste in un gruppo di malattie che comportano un coagulamento del
sangue non normale, provocando spesso la morte per emorragia, legata
all'incapacità di produrre una particolare proteina, Fattore VIII.
Questa malattia comportò in passato nella famiglie reali europee
molti morti, come alcuni figli di Elisabetta I d'Inghilterra e il
figlio dell'ultimo zar di Russia Nicola Romanov, Aleksej.
 |
| Fattore VIII |
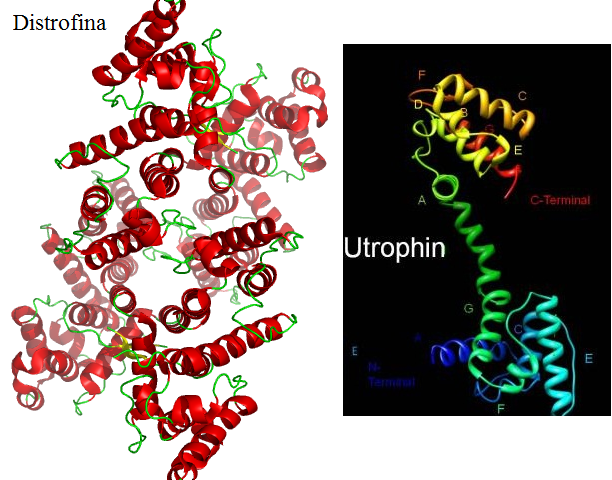
Un'altra malattia
legata al sesso è la distrofia muscolare di Duchenne che provoca una
grave insufficienza dei muscoli volontari. Ciò è dovuto da una
proteina chiamata distrofina che è presente in forma difettosa,
scoperta nel 1987 dal genetista Louis M. Kunkel. Per sconfiggere
questa malattia si possono impiantare cellule staminali che
permettono la rigenerazione del tessuto muscolare, oppure usando
farmaci che contengono l'utrofina, una proteina che sostituisce la
distrofina.

Un'altra malattia
importante è il favismo, dovuta alla carenza dell'enzima
glucosio-6-fosfato-deidrogenasi, presente nei globuli rossi, i quali
si autodistruggono (emolisi), provocando una forte anemia con
collasso cardiocircolatorio. Ciò è dovuto all'ingerimento di alcuni
cibi come le fave o alcuni farmaci che inibiscono la corretta
funzione di questo enzima.
 |
| Fave fresche |
 |
| Glucosio-6-fosfato deidrogenasi |
Infine la
sindrome dell'X fragile è anche molto colpita.



.jpg)




















.jpg)